Cirrhosis-associated immune dysfunction (CAID) – causes, phenotypes and consequences
Daniel Ján Havaj1, Ľubomír Skladaný1
+ Affiliation
Summary
The term cirrhosis-associated immune dysfunction represents a wide spectrum of alterations in the local and systematic immune response. CAID is characterised by parallel ongoing systematic inflammation and immune deficiency, the intensity of manifestation of which depends on the stage of the disease, predisposing and precipitating factors. The gut-liver axis plays a key role in the pathogenesis of CAID. Impaired interplay between the gut and liver and changes in microbiome composition are responsible for damage to the intestinal barrier and continuous antigen stimulation, leading to the new proinflammatory setting of the immune system, which represents the low-grade inflammation phenotype CAID. Episodic aggravation of microbial translocation and episodic effect of precipitating factors cause a burst of systematic inflammation and breakthrough of the immune tolerance. The result is dysregulated hyperinflammatory immune response, which represents the high-grade inflammation phenotype of CAID. The change of the phenotype stays in the background of transition to decompensated cirrhosis. The systemic inflammation increases across the subtypes of decompensation with the highest degree reached in the ACLF, which represents the fulminant immunophenotype of CAID. Hyperinflammatory immune response, immunopathologic and immunometabolic changes act synergistically with organ-specific mechanisms in the development of organ dysfunctions. Lasting dysregulated pro-inflammatory reaction leads to immunity exhaustion and innate and adaptive immune cells reprogramming. This immune paralysis is the cause of problematic infections and represents the indolent immunophenotype of CAID.
Keywords
cirrhosis-associated immune dysfunction (CAID), low-grade inflammation phenotype, high-grade inflammation phenotype, systematic inflammation, fulminant immunophenotype, immune paralysis, indolent immunophenotype, gut-liver axis, microbiome, inflammation
Introduction
Inflammation is an essential component in the process of tissue regeneration and is the critical pathophysiological pathway involved in the natural history of advanced chronic liver disease (ACLD), leading to cirrhosis and complications thereof. An aetiological cause of ACLD and dysbiosis drives the inflammatory reaction to restore homeostasis and activates the liver resident immune system to control the local response and prevent the inception of systematic inflammation. The consequence of a long-lasting cause of a particular aetiological factor is dysregulated immune response and subsequent chronic systematic inflammation with permanent release of proinflammatory mediators [1,2]. This process activates hepatic stellate cells and promotes fibrogenesis, leading to a distortion of the hepatic microarchitecture. This remodelling increases resistance to portal blood flow and results in the structural component of portal hypertension [3]. These changes correspond to the transition from chronic liver disease to cirrhosis.
Resident liver immunology
The unique arrangement of the liver parenchyma is responsible for metabolic and detoxicant function and is important for frontline immune response. Specialised innate and adaptive immune cells constitute about one-third of the total cell population in the liver. They comprise the carefully controlled network challenged by the continuous inflow of antigens from the gut or altered liver parenchyma. This microenvironment provides detection and subsequently clearance of pathogens and simultaneously maintains the balance between immunity and tolerance [4–6].
Kupffer cells are specialised liver resident macrophages mostly localised within the sinusoidal space. They are responsible for removing and degrading immunogenic molecules by phagocytosis, antigen presentation to T cells by MHC and costimulatory molecules, and the secretion of cytokines, chemokines and other inflammatory mediators. On the other hand, they induce immune tolerance [7].
Liver sinusoidal endothelial cells (LSEC) are highly specialised, fenestrated endothelial cells lining the hepatic sinusoids and they constitute the interface between blood cells, hepatocytes and hepatic stellate cells. This position is crucial to nutrient exchange, clearance and intrahepatic vascular tone regulation. LSECs represent gatekeepers for hepatic immunity because they function as antigen-presenting cells (APC), regulate leukocyte recruitment and in connection with PD-L1 expression, suppress the immune response and maintain immune tolerance by the generation of the anergic-like phenotype of an immune response [8,9].
Dendritic cells are derived from monocyte-macrophage progenitors residing around the central vein and periportal tracts. They sense the liver environment and after activation by antigen, they mature toward several functional antigens presenting cells (APCs) linking the innate and adaptive arms of the immune system. Dendritic cells are responsible for maintaining liver homeostasis and tolerance. They express MHC molecules and costimulatory molecules to present antigens to T-cells in a tolerogenic way. They promote the deletion of activated T-cells and activation of regulatory T-cells owing to the effect of their immuno-active products such as IL1, TGF-ß, CTLA-4, PD-1; they reduce the expression of TLR and crosstalk with NK and NKT cells. If the liver microenvironment changes its character to proinflammatory, dendritic cells switch their phenotype and contribute to the production of pro-inflammatory cytokines like TNF-α and IL6, provoking oxidative stress and the activation of stellate cells [10].
Lymphocytes play an important role in maintaining liver homeostasis. Innate lymphoid cells (NK cells, ILC 1, 2, 3s) and innate-like T-lymphocytes (NK cells, γδ T-cells, and MAITs) are scattered through the liver parenchyma and they share similar phenotypes and functional properties depending on the setting of the liver microenvironment. Many of them show tissue-resident features and act as sentinels and frontline defenders in response to antigen stimulation and function in immunosurveillance. They also contribute to immune regulation and maintain a tolerogenic environment [11,12].
Gut, microbiome and the gut-liver axis
In addition to digestion and absorption, the gut plays an important role in human immunity. This function is provided by gut-associated lymphoid tissue (GALT), the largest immunological organ in the human body. GALT is composed of diffuse lymphoid tissue, intestinal lymphoid follicles, Payers patches and mesenteric lymph nodes (MLNs). Effector immune cells of diffuse lymphoid tissue are distributed along the mucosal epithelium and lamina propria and represent the first defense against gut-derived antigens and pathogens [13,14].
Activated B cells differentiate into plasma cells and produce high levels of specific IgA antibodies responsible for the neutralisation of bacterial toxins and pathogens and healthy microbiota composition.
Intraepithelial and diffuse T-lymphocytes comprise two subtypes according to their phenotype and maturation process – conventional, which provide the defence against pathogens crossing the epithelial barrier, and unconventional, which have regulation functions and represent immune memory. The other component of this milieu is innate immune cells [15].
Innate lymphoid cells, such as NK, ILC 1–3 and LTi cells, can activate independently of the MHC complex. This is possible owing to a wide spectrum of surface receptors, which intermediate the rapid secretion of cytokines, prompt immune or tolerogenic response and crosstalk with other immune cells [16].
Macrophages are diffusely scattered throughout the intestinal wall and with transepithelial dendrites, they are in direct contact with the epithelial layer, double-layered mucus and intraluminal microbiome and metabolome content. This organisation enables them to capture and destroy in the process of phagocytosis any pathogens that breach the barrier, clearing impaired epithelial cells and they contribute to the regeneration of the epithelial barrier. Macrophages interplay with microbiota and in the physiological condition, they are adjusted to maintain gut homeostasis and immune tolerance through the production of immunoregulatory cytokines (IL-10, TGF-ß, etc). They also drive the maturation of Treg cells, Th17 lymphocytes and ILC3. Moreover, they transfer acquired antigens to dendritic cells, which are professionals responsible for the antigen presentation [17].
Dendritic cells are localised in lamina propria and they are responsible for transporting antigen to MLNs for presentation and modulation of the adaptive T-cell response. They also affect the maturation of B-cells and T-cells to effector intestine cells [18,19]. Lymphoid follicles and their aggregates, called Payers patches, act as immune sensors and play a role in immune response development. They integrate antigenic stimuli from the gut by reaching up into the follicles where they communicate with specialised cells (M cells), antigen-presenting cells, macrophages and dendritic cells, and create conditions for B-cell maturation and class switching with the help of T-lymphocytes. These cells then pass to the mesenteric lymph nodes, where the immune response is amplified [14].
Gut-liver axis. Between the gut, its microbiota, and the liver is a bidirectional relationship. Liver products through the biliary tract influence the composition of the gut microbiome and the permeability of the gut barrier. On the other hand, signals from the gut generated by dietary, genetic and environmental factors impact the liver through the portal vein, immunological, synthetic and metabolic signals. This interplay, called the gut-liver axis, has a key role in maintaining immune homeostasis and its disruption contributes to the pathogenesis and progression of liver diseases [20].
Low-grade inflammation as a driver of progression of liver disease
The liver resident immune system is incessantly exposed to low levels of antigens derived from the gut (called MAMPs or PAMPs [Microbial-associated molecular patterns and Pathogen-associated molecular patterns]) and damaged liver parenchyma (called DAMPs [Damage- -associated microbial patterns]). Because of highly specialised self-regulatory mechanisms mediated via TLR-receptors, these antigens rapidly induce and later resolve inflammatory responses to maintain homeostasis [1].
Pathogen-associated molecular patterns represent microbes or microbial components such as lipopolysaccharides (LPS), flagellin, lipoteichoic acid, peptidoglycan, bacterial-associated unmethylated CpG motifs, single- or double-stranded RNA or DNA, fungal beta-glucan, cytolysin, candidalysin, etc [4,21,22].
Damage-associated molecular patterns represent intracellular molecules released from damaged cells, such as nuclear and mitochondrial DNA, HMBG1, ATP, etc [23]. These distinctive molecules bind to specific pattern recognition receptors (PRRs) located on the membrane (Toll-like receptors, TLRs) or inside the immune cells (Nucleotide-binding oligomerisation domain-like receptors, NLRs, Retinoic acid-inducible gene I-like receptors, RIGRs) and consequently activate signalling pathways leading to an inflammatory response. This process activates the formation of the cytosolic multiprotein complex called inflammasome, which promotes transcription, maturation and secretion of proinflammatory cytokines such as IL-1ß and IL18 (canonical activation) or IL-1ß and IL-1α (non-canonical activation) [24,25]. Activated hepatocytes, Kupffer cells, macrophages and other resident myeloid cells release cytokines, which promote local and systematic inflammatory response, activation and recruitment of immune effector cells and further cytokine production. Also activated are resident T-cells to functional effector T-cells responsible for pathogen clearance. The first immune cells that infiltrate the liver are neutrophils that remove pathogens by phagocytosis, releasing antimicrobial agents and generating extracellular traps. Simultaneously, neutrophils release a large number of proinflammatory cytokines (TNF-α, IL-1b, IL-6, plate-derived growth factor, TGF-ß) and express activation markers (CD11b and CD62L). Other recruited cells are monocytes, which differentiate into macrophages and change their activation state and phenotype to M1 – proinflammatory or M2 – anti-inflammatory. Released cytokines also activate hepatic stellate cells responsible for extracellular matrix production and fibrogenesis initiation [6,26]. Physiologically, for the resolution of fibrosis, the responsible cells are macrophages, NK cells, and CD8+ tissue-resident memory T [27].
Persistent antigen stimulation from the gut together with an underlying liver disease generates a continuous pro-inflammatory response that overcomes anti-inflammatory mechanisms. This process comprises intracellular changes leading to abnormal sensitivity of signalling pathways to lipopolysaccharide, defective production of the anti-inflammatory cytokines such as IL-10 by Kupffer cells and LSECs [28], monocyte and macrophage polarisation toward M1 pro-inflammatory phenotype persistently releasing proinflammatory cytokines such as TNF-α, IL-1b, IL-6, etc [29]. These changes lead to progressive loss of tolerance and an excessive pro-inflammatory response upon antigen recognition. Chronic inflammation is responsible for progressive fibrogenesis and architectural distortion of the liver and contributes to haemodynamic disturbances and the development of portal hypertension [26,30].
An anatomical and functional relationship between the gut and the liver plays a key role in the pathogenesis of cirrhosis and the development of complications of cirrhosis [31–33]. The microbiome composition and intestinal functions are affected by several aetiological factors of chronic liver diseases, such as alcohol use and a diet low in fiber, and on the other hand by changes during the progression of chronic liver disease [34,35]. Decreased gut motility caused by activated compensatory neurohumoral mechanism, reduced primary and secondary bile acid synthesis, decreased enterohepatic circulation, impaired FXR signalling which is important in the epithelial barrier, and portal hypertension contribute to gut and microbiome changes [13].
The shift in the microbiome composition comprises reduced bacterial diversity (number of different species and composition of species), imbalance in potentially pathogenic bacteria (Fusobacteria, Proteobacteria, Enterococcaceae and Streptococacceae), and potentially beneficial autochthonous bacteria (Bacteroidetes, Ruminococcus, Roseburia, Veillonellaceae and Lachnospiraceae), increased bacterial load and bacterial overgrowth. Alterations of the gut microbiome (dysbiosis) are comprised not only by bacteriome but also fungome and virome [36–39]. Bacterial overgrowth is associated with decreased motility and increased adherence of bacteria to the mucosal and epithelial layer, which results in fermentation of the luminal content and changes in the microbial metabolites, leading to thinning of the mucosal layer and loosening of epithelial tight junctions. Leaky gut ensues with a disruption of the intestinal barrier, leading to activation of mucosal immune cells and bacterial translocation to the portal and systematic circulation [40–43]
Dysbiosis, bacterial overgrowth, increased adherence to the epithelial layer, breakdown of the intestinal-vascular barrier and bacterial translocation lead to persistent stimulation of gut-associated lymphatic tissue, resulting in subclinical inflammation. These changes are evidenced by the higher number of activated pro-inflammatory monocytes and dendritic cells, the higher number of T-lymphocytes polarised to Th1 regulatory phenotype and concomitant Th17 depletion [44,45]. The inflammatory process decreases secretion of antibacterial peptides, IgA, defensins, which along with pro-inflammatory cytokines such as IL6, TNF and INF gamma boost bacterial overgrowth and translocation [20,46–48].
The gut thus becomes the driver of low-grade systemic and liver inflammation and chronic liver disease progression. Microbial components and viable bacteria reach the portal circulation and as PAMPs activate inflammatory response in the liver. In addition, some of the bacterial products such as cytolysin (secreted by Enterococcus faecalis) and candidalysin (secreted by Candida albicans) translocated from the gut, directly damage hepatocytes and activate inflammatory response through DAMPs [13,49]. The inflammatory reaction in the liver is augmented by proinflammatory cytokines and activated immune cells released from the GALT. The progression of liver disease is also facilitated by proangiogenic molecules liberated from GALT, that promote angiogenesis and contribute to the development of portal hypertension [50–52]. This proinflammatory setting of the immune system represents the low-grade systematic inflammatory phenotype of CAIDS [4].
High-grade inflammation as the driver of decompensation and ACLF
Local inflammation in the liver driven by the proinflammatory setting of the gut-liver axis leads to the progression of chronic liver disease to cirrhosis. The breakthrough in immune tolerance is responsible for dysregulated immune response and the development of systematic inflammation. [53,54] This change is displayed by increased plasma levels of acute-phase proteins such as CRP, LPS-binding protein, pro-inflammatory cytokines (TNF, IL-1ß, IL-6, IL-8, IL-17, IFNγ) and their soluble receptors such as TNF soluble receptors I and II, and as cirrhosis, progresses to decompensated or ACLF continuous increase anti-inflammatory cytokines. (IL-10, IL-1RA) [55] The profile of circulating cytokines depends particularly on the aetiology of cirrhosis: while in alcohol-associated liver disease (ALD) IL-8 dominates, in chronic hepatitis B and C IL-6 and IL-18 dominate [56]. Lasting recruitments of immune cells into the liver represent an increased level of chemokines and endothelial activation markers (MIP, P-selectin, ICAM1, VCAM1, VEGF, etc). Inflammatory cytokines activate circulating immune cells [57]. Activated neutrophils increased their respiratory burst activity and express activation marker CD11b (receptor which mediates adhesion and cytotoxic activity) [58]. Circulating monocytes show an increased level and predominance of non-classical CD14+16+ pro-inflammatory and pro-fibrotic subset with increased expression of human leukocyte antigen DR (HLA-DR) and costimulatory molecules (CD80/CD86) and increased TNF production [59–61]. For activated T-cell, polarisation to TH1 – proinflammatory phenotype, which produces high levels of IFN-γ, is typical [62]. Another sign of B-cell activation is increased expression of HLA-DR and costimulatory molecules [63].
Parallel with systematic inflammation, portal hypertension progresses and specific compensatory mechanisms are activated: splanchnic vasodilatation, activation of the compensatory neurohumoral mechanisms, the hyperkinetic circulation and portosystemic shunting. Portal hypertension and systematic inflammation, which also aggravate the dynamic component of portal hypertension, are pivotal factors in the transition from compensated to decompensated cirrhosis and the development of ascites, hepatic encephalopathy and variceal haemorrhage [26]. The systemic inflammation increases across the subtypes of acute decompensation (stable decompensated cirrhosis SDC > unstable decompensated cirrhosis UDC > pre-ACLF), with the highest degree reached in the ACLF phenotype where it intermediates multiple organ dysfunction [64]. The reason for the burst of systematic inflammation is an episodic aggravation of translocation of viable bacteria and bacterial products, or precipitant events such as bacterial infection, alcoholic hepatitis, the flare of hepatitis, etc [65].
Systematic inflammation plays an important role in the organ dysfunction associated with decompensated cirrhosis and ACLF by its synergic effect with already established effective arterial hypovolemia, portal hypertension, hyperammonemia, immune pathology and metabolic dysregulation [66].
Proinflammatory cytokines stimulate endothelial cells in splanchnic circulation to produce NO and other vasodilators, which intensify splanchnic vasodilatation and effective blood volume reduction. These changes simultaneously hyperactivate the compensatory mechanism and increase the level of circulating vasoconstrictors, such as noradrenaline, renin-angiotensin-aldosterone axis and vasopressin. [26,67,68].
Haemodynamic changes and exposure to vasoactive and pro-inflammatory substances induce and aggravate cirrhotic cardiomyopathy and cardiovascular dysfunction [69,70]. Proinflammatory cytokines also activate hepatic-stellate cells, which increase their fibroproduction and contractibility, and hand in hand with the local imbalance of vasoactive substances they significantly increase a dynamic component of portal hypertension [71]. Inflammatory molecules furthermore affect tissue and somatic cells and induce their damage in a process called immunopathology [66].
In the kidneys, they affect the highly metabolic active part – tubular cells – and cause mitochondrial injury, upregulation of renal TLR, inflammasome activation, hypersecretion of TNF alpha into the luminal fluid, leukocyte infiltration and oxidative burst damage, leading to tubular damage. In addition, they induce endothelial dysfunction, capillary micro-thrombosis and ischaemic injury. All these changes aggravate haemodynamically induced renal dysfunction and contribute to HRS-AKI (hepatorenal syndrome of the acute kidney injury type) [72–75].
Proinflammatory cytokines also affect the brain tissue and, along with hyperammonemia and translocated gut-driven microbial products and inflammatory substances, participate in the development of hepatic encephalopathy. They activate endothelial and resident immune cells such as microglia and astrocytes, induce local cytokine production and disrupt neurotransmission. Simultaneously, they affect the permeability of the haematoencephalic barrier, leading to the recruitment of circulating immune cells to the brain and a higher supply of ammonia and other toxic substances. These changes aggravate the clinical manifestation of hepatic encephalopathy [76].
Systematic inflammation is responsible for immunometabolic changes because of high metabolic demand requiring the reallocation of nutrients. Proinflammatory cytokines stimulate the hypothalamic-pituitary-adrenal axis and the sympathetic nervous system to proteo- and lipolysis, subsequently leading to the release of amino acids and lipids to fuel hypermetabolic active immune cells. In parallel, lipids and their derivates, acting directly as immune stimulants and amino acids (and ammonia) worsen mitochondrial function, leading to catabolism and neutrophil dysfunction. These cytokines also inhibit nutrient consumption in peripheral tissue. These processes lead to catabolism and loss of skeletal muscle mass and function (sarcopenia) and adipose tissue mass [77]. In addition, cytokines cause the metabolic switch to reduced oxidative glucose metabolism in the mitochondria, increased extramitochondrial glucose utilisation through aerobic glycolysis and pentose-phosphate pathway leading to decreased ATP production and increased ROS production responsible for further tissue injury and DAMPs release. Immunometabolic changes play an important role in the disease progression and progressive exhaustion of the immune system [26].
These changes represent a high-grade inflammation phenotype of CAIDS. [4] Systemic inflammation is the major driver of progression from compensated to decompensated cirrhosis, acting synergistically with organ-specific mechanisms in the development of major complications of cirrhosis and there is a specific connection to mortality. ACLF, a syndrome characterised by acute decompensation of chronic liver disease which is associated with organ failure and high short-term mortality, represents an example of the most severe systemic inflammation (e. g. much higher than in sepsis) [55,68]. This is called the fulminant immunophenotype of CAIDS and is exemplified by elevated CRP, white blood cells, and pro-inflammatory cytokines, together with increased anti-inflammatory cytokines and soluble markers of macrophage activation (sCD163 and mannose receptor) [78]. Discerning this immunological subtype of ACLF could lead to personalised prognostic stratification and therapy – e. g. the plasma exchange (Fig. 1) [79].
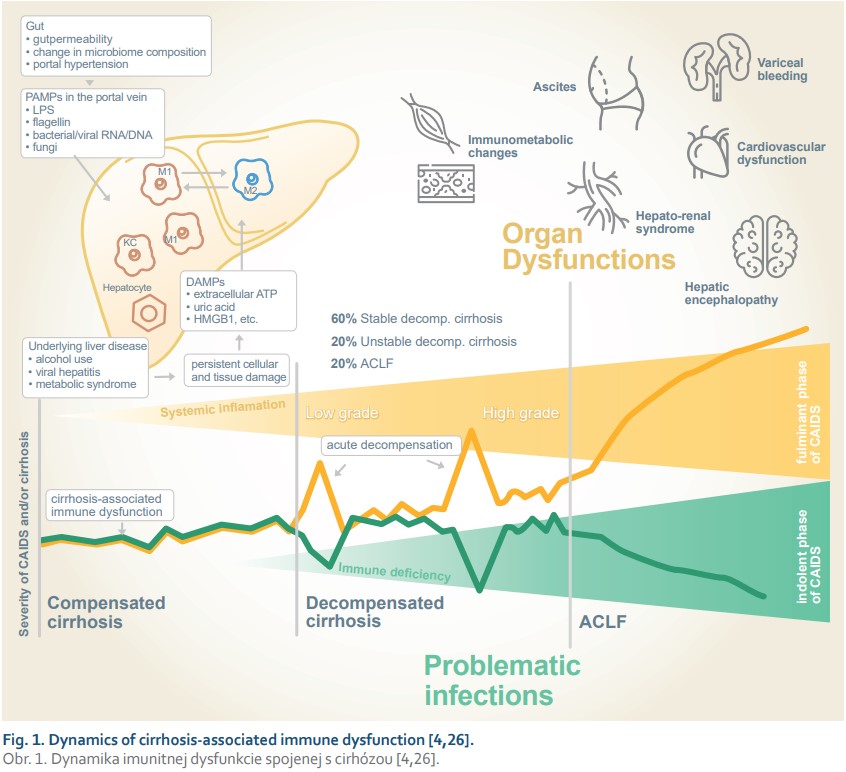
Immune paralysis
Chronic systemic inflammation with dysregulated pro-inflammatory reactions and damage of barriers in diverse compartments all cause immune cell exhaustion and reprogramming. Continuously stimulated immune cells switch into non-responsive phenotype leading to dysfunctional effector response and immune paresis. Whilst each cirrhosis aetiology has its distinctive features of immune paresis, most of the pathophysiological mechanisms are common to all aetiologies [66,79].
Innate immune cells
Neutrophils, thought of as pawns of the immune system, are paralysed in many of their functions [60,80,81]. Long-lasting exposition to proinflammatory cytokines affects many intracellular pathways leading to defect chemotaxis, migration (reduced expression CXCR1 and CXCR2 receptors [82], defect activation IL33/ST2 [83]), opsonisation, impaired phagocytic activity, respiratory burst (defect activation AKT, MAPK, NADPH oxidase pathway) [84,85] and reduced formation of neutrophil extracellular traps [86–88]. Besides functional derangement to immune paresis contributes also splenic sequestration [89].
Monocytes. The switch to the immunosuppressed state also impacts monocytes. [60] Immunogenetic and immunometabolic reprogramming of mainly classical (CD14+CD16−) monocytes is responsible for defective chemotaxis, phagocytosis (e. g. Fcγ receptor impairment) [90], enzymatic and superoxide production (dampened expression of IRIF and PRKCE is associated with defective NADPH oxidase) [91]. Abundantly present in the circulation are immunoregulatory monocytes − AXL-expressing monocyte population (CD14+CD16highHLA-DRhigh) characterised by attenuated TNF-α/IL-6 responses and T-cell activation [92] and monocytes that express MER receptor tyrosine kinase (MERTK) responsible for downregulation of innate inflammatory immune responses and inhibition of TLR activation and pro-inflammatory cytokine production [93]. Functional analyses of immunoparalysed monocytes show decreased HLA-DR expression, which refers reduced antigen-presenting ability, decreased production of pro-inflammatory cytokines (e. g. IFN-γ, TNF-α, IL-6) after microbial challenge through NF-kB pathway inhibition, and enhanced anti-inflammatory (e. g. SLPI) mediator secretion [94–96].
Adaptive immune cells
Ineffective production of naïve T-cells because of defective thymopoiesis [97] and increased splenic sequestration is responsible for T-cell depletion in circulation. T-cell lymphopenia is also worsened by an imbalance between apoptosis and peripheral proliferation. Continuously stimulated memory lymphocytes express a higher level of apoptosis marker CD95+, and they are unable to proliferate in response to new antigenic stimulation [98]. In circulation, dysregulated specific types of lymphocytes are also present. MAIT cells (mucosal-associated invariant T-cells) with high antimicrobial potency are reduced, hyperactivated and incompetent in response to antimicrobial and cytokine stimulation [99,100]. Also present is the immunosuppressive subset of HLA-DR+CD8+T-cells with co-expression PD1 (program cell death protein 1), TIM3 (T-cell immunoglobulin and mucin domain-containing protein 3) resulting in reduced TNF and IFN production [101,102]. The B-cell compartment is also affected due to reduced count and pathological function, especially in the memory cell subset. The loss of CD27+ memory cells associated with impaired TNF-ß and IgG production and impaired allostimulatory capacity contributes to immune paresis. The remaining B-cells are less effective at stimulating CD4+ T-cell responses, hyporesponsive to activation via CD40 and TLR9, with impaired upregulation of costimulatory markers, production of TNF-ß and production of IgG [103]. Dysfunction of NK cells with reduced cytotoxic activity also contributes to immune paralysis [104].
Lasting systematic inflammation triggers the expansion of mononuclear CD14+CD15−CD11b+HLA-DR− myeloid-derived suppressor cells (M-MDSCs), which continuously dampen the immune response. M-MDSCs are responsible for suppressing T-cell activation, pathogen uptake and Toll-like receptor (TLR) -elicited proinflammatory responses to microbial challenge. They are associated with ACLF, and their persistence during disease progression predicts a poor outcome and an increased incidence of infections [105].
A key role in immune reprogramming is also played by soluble molecules such as anti-inflammatory cytokines IL-10 and IL-1RA. IL-10 through inhibition of NF-kB reduces monocyte secretion of TNF, IL-1, IL-6, IL-8 and IL-12, reduces the secretion of IFN-γ and correlates with the expression of inhibitory receptors in monocytes (MERKT), lymphocytes (PD1) and TIM3 [106–108].
In addition to changes in immune cells and circulating immune molecules, structural derangements of liver parenchyma contribute to the immunological burn-out. Capillarisation of sinusoids, fibrosis and a damaged reticuloendothelial system cause reduced decreased clearance, portosystemic shunting leads to the continual spread of microbes and distinctive molecules to systematic circulation, impairment of Kupfer cells contribute to immune dysregulation [109]. Moreover, decreasing the synthesis of immune proteins and receptors such as complement, soluble PRRs, albumin, proteins of the acute phase (CRP, MBL, hepcidin, fibrinogen, proteinase inhibitors) plays a role in an incompetent immune response [110]. Albumin is physiologically able to bind immunoactive molecules (pro-inflammatory and immunosuppressive) such as prostaglandin E (PGE2). Because of structural dysfunction and decreased concentration of albumin, the plasmatic concentration of PGE2 is significantly higher and it contributes to macrophage dysfunction [111,112]. Other molecules that have a negative impact on immune tolerance are circulating catecholamines, tryptophan catabolites and ammonia, causing neutrophil swelling and impaired phagocytosis [113].
Conclusion
Dynamic alterations of the immune system which parallel the pathophysiological cascade from chronic liver disease to liver cirrhosis to ACLF are comprised of the spectrum of phenotypes of CAIDS from long-lasting low-grade inflammation of CLD to a high-grade inflammation of acute decompensation of cirrhosis and ACLF, to immune exhaustion and so-called indolent death. The disruption of tolerogenic mechanisms in the liver leads to low-grade systematic inflammation, which is responsible for progressive fibrogenesis. A key role in the disease progression is played by the gut-liver axis, because of continuous antigen stimulation driving inflammatory response and cirrhosis progression. The ongoing dysregulation of systematic immune response leads to high-grade inflammation, which acts complementarily to organ-specific mechanisms in the development of cirrhosis decompensation or ACLF and a fulminant phase of CAIDS. Long-lasting high-grade inflammation causes immune cell exhaustion and reprogramming to non-responsive phenotype – an indolent immunophenotype of CAIDS. This phase is characterised by problematic infections by opportunist pathogens [114–116]. It is very important to recognise these phenotypes, as they represent the evolutionary phases of progression, convey prognosis and govern the management.
ORCID authors
D. J. Havaj ORCID 0000-0001-5979-8326,
Ľ. Skladaný ORCID 0000-0001-5171-3623.
Doručené/Submitted: 16. 3. 2022
Prijaté/Accepted: 28. 3. 2022
Daniel Ján Havaj, MD
Internal Med Department II
Slovak Medical University
Division of Hepatology, Gastroenterology and Liver Transplantation
FD Roosevelt Hospital
L. Svobodu 1 Square
975 17 Banska Bystrica
havaj.daniel@gmail.com
To read this article in full, please register for free on this website.
Benefits for subscribers
Benefits for logged users
Literature
1. Dirchwolf M, Ruf AE. Role of systemic inflammation in cirrhosis: from pathogenesis to prognosis. World J Hepatol 2015; 7(16): 1974–1981. doi: 10.4254/wjh.v7.i16.1974.
2. Zhou WC, Zhang QB, Qiao L. Pathogenesis of liver cirrhosis. World J Gastroenterol 2014; 20(23): 7312–7324. doi: 10.3748/wjg.v20.i23.7312.
3. Novo E, Bocca C, Foglia B et al. Liver fibrogenesis: un update on established and emerging basic concepts. Arch Biochem Biophys 2020; 689: 108445. doi: 10.1016/j.abb.2020.108445.
4. Albillos A, Martin-Mateos R, Van der Merwe S et al. Cirrhosis-associated immune dysfunction. Nat Rev Gastroenterol Hepatol 2022; 19(2): 112–134. doi: 10.1038/s41575-021-00520-7.
5. Racanelli V, Rehermann B. The liver as an immunological organ. Hepatology 2006; 43(2 Suppl 1): S54–62. doi: 10.1002/hep.21060.
6. Robinson M, Harmon C, O’Farrelly C. Liver immunology and its role in inflammation and homeostasis. Cell Mol Immunol 2016; 13(3): 267–276. doi: 10.1038/cmi.2016.3.
7. Nguyen-Lefebvre AT, Horuzsko A. Kupffer cell metabolism and function. J Enzymol Metab 2015; 1(1): 101.
8. Shetty S, Lalor PF, Adams DH. Liver sinusoidal endothelial cells – gatekeepers of hepatic immunity. Nat Rev Gastroenterol Hepatol 2018; 15(9): 555–567. doi: 10.1038/s41575-018-0020-y.
9. Wilkinson AL, Qurashi M, Shetty S. The role of sinusoidal endothelial cells in the axis of inflammation and cancer within the liver. Front Physiol 2020; 11: 990. doi: 10.3389/fphys.2020.00990.
10. Méndez-Sánchez N, Córdova-Gallardo J, Barranco-Fragoso B et al. Hepatic dendritic cells in the development and progression of metabolic steatohepatitis. Front Immunol 2021; 12: 641240. doi: 10.3389/fimmu.2021.641240.
11. Wang Y, Zhang C. The roles of liver-resident lymphocytes in liver diseases. Front Immunol 2019; 10: 1582. doi: 10.3389/fimmu.2019.01582.
12. Jin-Seok Byun, Hyon-Seung Yi. Hepatic immune microenvironment in alcoholic and nonalcoholic liver disease. Biomed Res Int 2017; 2017: 6862439. doi: 10.1155/2017/6862439.
13. Albillos A, de Gottardi A, Rescigno M. The gut-liver axis in liver disease: pathophysiological basis for therapy. J Hepatol 2020; 72(3): 558–577. doi: 10.1016/j.jhep.2019.10.003.
14. Mörbe UM, Jørgensen PB, Fenton TM et al. Human gut-associated lymphoid tissues (GALT); diversity, structure, and function. Mucosal Immunol 2021; 14(4): 793–802. doi: 10.1038/s41385-021-00389-4.
15. Ma H, Tao W, Zhu S. T lymphocytes in the intestinal mucosa: defense and tolerance. Cell Mol Immunol 2019; 16(3): 216–224. doi: 10.1038/s41423-019-0208-2.
16. Jiao Y, Wu L, Huntington ND et al. Crosstalk between gut microbiota and innate immunity and its implication in autoimmune diseases. Front Immunol 2020; 11: 282. doi: 10.3389/fimmu.2020.00282.
17. Bain CC, Schridde A. Origin, differentiation, and function of intestinal macrophages. Front Immunol 2018; 9: 2733. doi: 10.3389/fimmu.2018.02733.
18. Schiavi E, Smolinska S, O’Mahony L. Intestinal dendritic cells. Curr Opin Gastroenterol 2015; 31(2): 98–103. doi: 10.1097/mog.0000 000000000155.
19. Faria AMC, Reis BS, Mucida D. Tissue adaptation: implications for gut immunity and tolerance. J Exp Med 2017; 214(5): 1211–1226. doi: 10.1084/jem.20162014.
20. Trebicka J, Macnaughtan J, Schnabl B et al. The microbiota in cirrhosis and its role in hepatic decompensation. J Hepatol 2021; 75(Suppl 1): S67–S81. doi: 10.1016/j.jhep.2020.11.013.
21. Mogensen TH. Pathogen recognition and inflammatory signaling in innate immune defenses. Clin Microbiol Rev 2009; 22(2): 240–273. doi: 10.1128/CMR.00046-08.
22. Chen P, Stärkel P, Turner JR et al. Dysbiosis-induced intestinal inflammation activates tumor necrosis factor receptor I and mediates alcoholic liver disease in mice. Hepatology 2015; 61(3): 883–894. doi: 10.1002/hep.27489.
23. Kubes P, Mehal WZ. Sterile inflammation in the liver. Gastroenterology 2012; 143(5): 1158–1172. doi: 10.1053/j.gastro.2012.09.008.
24. Kelley N, Jeltema D, Duan Y et al. The NLRP3 inflammasome: an overview of mechanisms of activation and regulation. Int J Mol Sci 2019; 20(13): 3328. doi: 10.3390/ijms20133328.
25. Wu X, Dong L, Lin X et al. Relevance of the NLRP3 inflammasome in the pathogenesis of chronic liver disease. Front Immunol 2017; 8: 1728. doi: 10.3389/fimmu.2017.01728.
26. Engelmann C, Clària J, Szabo G et al. Pathophysiology of decompensated cirrhosis: portal hypertension, circulatory dysfunction, inflammation, metabolism and mitochondrial dysfunction. J Hepatol 2021; 75(Suppl 1): S49–S66. doi: 10.1016/j.jhep.2021.01.002.
27. Koda Y, Teratani T, Chu PS et al. CD8+ tissue-resident memory T cells promote liver fibrosis resolution by inducing apoptosis of hepatic stellate cells. Nat Commun 2021; 12(1): 4474. doi: 10.1038/s41467-021-24734-0.
28. Heymann F, Peusquens J, Ludwig-Portugall I et al. Liver inflammation abrogates immunological tolerance induced by Kupffer cells. Hepatology 2015; 62(1): 279–291. doi: 10.1002/hep.27793.
29. Wang C, Ma C, Gong L et al. Macrophage polarization and its role in liver disease. Front Immunol 2021; 12: 803037. doi: 10.3389/fimmu. 2021.8030.
30. Noor MT, Manoria P. Immune dysfunction in cirrhosis. J Clin Transl Hepatol 2017; 5(1): 50–58. doi: 10.14218/JCTH.2016.00056.
31. Trebicka J, Bork P, Krag A et al. Utilizing the gut microbiome in decompensated cirrhosis and acute-on-chronic liver failure. Nat Rev Gastroenterol Hepatol 2021; 18(3): 167–180. doi: 10.1038/s41575-020-00376-3.
32. Almeida JI, Tenreiro MF, Martinez-Santamaria L et al. Hallmarks of the human intestinal microbiome on liver maturation and function. J Hepatol 2022; 76(3): 694–725. doi: 10.1016/j.jhep.2021.10.015.
33. Tripathi A, Debelius J, Brenner DA et al. The gut-liver axis and the intersection with the microbiome. Nat Rev Gastroenterol Hepatol 2018; 15(7): 397–411. doi: 10.1038/s41575-018-0011-z.
34. Bajaj JS. Alcohol, liver disease and the gut microbiota. Nat Rev Gastroenterol Hepatol 2019; 16(4): 235–246. doi: 10.1038/s41575-018-0099-1.
35. Bajaj JS, Idilman R, Mabudian L et al. Diet affects gut microbiota and modulates hospitalization risk differentially in an international cirrhosis cohort. Hepatology 2018; 68(1): 234–247. doi: 10.1002/hep.29791.
36. Solé C, Guilly S, Da Silva K et al. Alterations in gut microbiome in cirrhosis as assessed by quantitative metagenomics: relationship with acute-on-chronic liver failure and prognosis. Gastroenterology 2021; 160(1): 206–218. doi: 10.1053/j.gastro.2020.08.054.
37. Bajaj JS, Heuman DM, Hylemon PB et al. Altered profile of human gut microbiome is associated with cirrhosis and its complications. J Hepatol 2014; 60(5): 940–947. doi: 10.1016/j.jhep.2013.12.019.
38. Qin N, Yang F, Li A et al. Alterations of the human gut microbiome in liver cirrhosis. Nature 2014; 513(7516): 59–64. doi: 10.1038/nature 13568.
39. Chen Y, Yang F, Lu H et al. Characterization of fecal microbial communities in patients with liver cirrhosis. Hepatology 2011; 54(2): 562–572. doi: 10.1002/hep.24423.
40. Pijls KE, Jonkers DMAE, Elamin EE et al. Intestinal epithelial barrier function in liver cirrhosis: an extensive review of the literature. Liver Int 2013; 33(10): 1457–1469. doi: 10.1111/liv.12271.
41. Muñoz L, Borrero MJ, Úbeda M et al. Intestinal immune dysregulation driven by dysbiosis promotes barrier disruption and bacterial translocation in rats with cirrhosis. Hepatology 2019; 70(3): 925–938. doi: 10.1002/hep.30349.
42. Assimakopoulos SF, Tsamandas AC, Tsiaoussis GI et al. Altered intestinal tight junctions’ expression in patients with liver cirrhosis: a pathogenetic mechanism of intestinal hyperpermeability. Eur J Clin Invest 2012; 42(4): 439–446. doi: 10.1111/j.1365-2362.2011.02 609.x.
43. Wiest R, Lawson M, Geuking M. Pathological bacterial translocation in liver cirrhosis. J Hepatol 2014; 60(1): 197–209. doi: 10.1016/ j.jhep.2013.07.044.
44. Muñoz L, Borrero MJ, Ubeda M et al. Interaction between intestinal dendritic cells and bacteria translocated from the gut in rats with cirrhosis. Hepatology 2012; 56(5): 1861–1869. doi: 10.1002/hep.25854.
45. Inamura T, Miura S, Tsuzuki Y et al. Alteration of intestinal intraepithelial lymphocytes and increased bacterial translocation in a murine model of cirrhosis. Immunol Lett 2003; 90(1): 3–11. doi: 10.1016/j.imlet.2003.05.002.
46. Kaliannan K. Compromise of alfa-defensin function in liver cirrhosis facilitates the toxic relationship between gut permeability and endotoxemia. Dig Dis Sci 2018; 63(10): 2492–2494. doi: 10.1007/s10620-018-5197-y.
47. Du Plessis J, Vanheel H, Janssen CEI et al. Activated intestinal macrophages in patients with cirrhosis release NO and IL-6 that may disrupt intestinal barrier function. J Hepatol 2013; 58(6): 1125–1132. doi: 10.1016/j.jhep.2013.01.038.
48. Genescà J, Martí R, Rojo F et al. Increased tumour necrosis factor alpha production in mesenteric lymph nodes of cirrhotic patients with ascites. Gut 2003; 52(7): 1054–1059. doi: 10.1136/gut.52.7.1054.
49. Wang R, Tang R, Li B et al. Gut microbiome, liver immunology, and liver diseases. Cell Mol Immunol 2021; 18(1): 4–17. doi: 10.1038/s41 423-020-00592-6.
50. Hassan M, Moghadamrad S, Sorribas M et al. Paneth cells promote angiogenesis and regulate portal hypertension in response to microbial signals. J Hepatol 2020; 73(3): 628–639. doi: 10.1016/j.jhep.2020.03.019.
51. Schierwagen R, Alvarez-Silva C, Madsen MSA et al. Circulating microbiome in blood of different circulatory compartments. Gut 2019; 68(3): 578–580. doi: 10.1136/gutjnl-2018-316227.
52. Alvarez-Silva C, Schierwagen R, Pohlmann A et al. Compartmentalization of immune response and microbial translocation in decompensated cirrhosis. Front Immunol 2019; 10: 69. doi: 10.3389/fimmu.2019.00069.
53. Albillos A, Lario M, Álvarez-Mon M. Cirrhosis-associated immune dysfunction: distinctive features and clinical relevance. J Hepatol 2014; 61(6): 1385–1396. doi: 10.1016/j.jhep.2014.08.010.
54. Martin-Mateos R, Alvarez-Mon M, Albillos A. Dysfunctional immune response in acute-on-chronic liver failure: it takes two to tango. Front Immunol 2019; 10: 973. doi: 10.3389/fimmu.2019.00973.
55. Clària J, Stauber RE, Coenraad MJ et al. Systemic inflammation in decompensated cirrhosis: characterization and role in acute-on-chronic liver failure. Hepatology 2016; 64(4): 1249–1264. doi: 10.1002/hep.28740.
56. Falasca K, Ucciferri C, Dalessandro M et al. Cytokine patterns correlate with liver damage in patients with chronic hepatitis B and C. Ann Clin Lab Sci 2006; 36(2): 144–150.
57. Martínez-Esparza M, Tristán-Manzano M, Ruiz-Alcaraz AJ et al. Inflammatory status in human hepatic cirrhosis. World J Gastroenterol 2015; 21(41): 11522–11541. doi: 10.3748/wjg.v21.i41.11522.
58. Liu K, Wang FS, Xu R. Neutrophils in liver diseases: pathogenesis and therapeutic targets. Cell Mol Immunol 2021; 18(1): 38–44. doi: 10.1038/s41423-020-00560-0.
59. Zimmermann HW, Seidler S, Nattermann J et al. Functional contribution of elevated circulating and hepatic non-classical CD14CD16 monocytes to inflammation and human liver fibrosis. PLoS One 2010; 5(6): e11049. doi: 10.1371/journal.pone.0011049.
60. Bernsmeier C, van der Merwe S, Périanin A. Innate immune cells in cirrhosis. J Hepatol 2020; 73(1): 186–201. doi: 10.1016/j.jhep.2020.03.027.
61. Irvine KM, Ratnasekera I, Powell EE et al. Causes and consequences of innate immune dysfunction in cirrhosis. Front Immunol 2019; 10: 293. doi: 10.3389/fimmu.2019.0029.
62. Muñoz L, Albillos A, Nieto M et al. Mesenteric Th1 polarization and monocyte TNF-alfa production: first steps to systemic inflammation in rats with cirrhosis. Hepatology 2005; 42(2): 411–419. doi: 10.1002/hep.20799.
63. Girón JA, Alvarez-Mon M, Menéndez-Caro JL et al. Increased spontaneous and lymphokine- conditioned IgA and IgG synthesis by B cells from alcoholic cirrhotic patients. Hepatology 1992; 16(3): 664–670. doi: 10.1002/hep.1840160309.
64. Trebicka J, Fernandez J, Papp M et al. The PREDICT study uncovers three clinical courses of acutely decompensated cirrhosis that have distinct pathophysiology. J Hepatol 2020; 73(4): 842–854. doi: 10.1016/j.jhep.2020.06.013.
65. Gustot T, Stadlbauer V, Laleman W et al. Transition to decompensation and acute-on-chronic liver failure: role of predisposing factors and precipitating events. J Hepatol 2021; 75(Suppl 1): S36–S48. doi: 10.1016/j.jhep.2020.12.005.
66. Arroyo V, Angeli P, Moreau R et al. The systemic inflammation hypothesis: towards a new paradigm of acute decompensation and multiorgan failure in cirrhosis. J Hepatol 2021; 74(3): 670–685. doi: 10.1016/j.jhep.2020.11.048.
67. Schrier RW, Arroyo V, Bernardi M et al. Peripheral arterial vasodilation hypothesis: a proposal for the initiation of renal sodium and water retention in cirrhosis. Hepatology 1988; 8(5): 1151–1157. doi: 10.1002/hep.1840080532.
68. Arroyo V. Pathophysiology, diagnosis and treatment of ascites in cirrhosis. Ann Hepatol 2002; 1(2): 72–79.
69. Møller S, Danielsen KV, Wiese S et al. An update on cirrhotic cardiomyopathy. Expert Rev Gastroenterol Hepatol 2019; 13(5): 497–505. doi: 10.1080/17474124.2019.1587293.
70. Sampaio F, Pimenta J. Left ventricular function assessment in cirrhosis: current methods and future directions. World J Gastroenterol 2016; 22(1): 112–125. doi: 10.3748/wjg.v22.i1.112.
71. Gracia-Sancho J, Marrone G, Fernández-Iglesias A. Hepatic microcirculation and mechanisms of portal hypertension. Nat Rev Gastroenterol Hepatol 2019; 16(4): 221–234. doi: 10.1038/s41575-018-0097-3.
72. Peerapornratana S, Manrique-Caballero CL, Gómez H et al. Acute kidney injury from sepsis: current concepts, epidemiology, pathophysiology, prevention and treatment. Kidney Int 2019; 96(5): 1083–1099. doi: 10.1016/j.kint.2019.05.026.
73. Gomez H, Ince C, De Backer D et al. A unified theory of sepsis-induced acute kidney injury. Shock 2014; 41(1): 3–11. doi: 10.1097/SHK.0000 000000000052.
74. Alobaidi R, Basu RK, Goldstein SL et al. Sepsis-associated acute kidney injury. Semin Nephrol 2015; 35(1): 2–11. doi: 10.1016/j.semnephrol.2015.01.002.
75. Angeli P, Garcia-Tsao G, Nadim MK et al. News in pathophysiology, definition and classification of hepatorenal syndrome: a step beyond the International Club of Ascites (ICA) consensus document. J Hepatol 2019; 71(4): 811–822. doi: 10.1016/j.jhep.2019.07.002.
76. Manzhalii E, Virchenko O, Falalyeyeva T et al. Hepatic encephalopathy aggravated by systemic inflammation. Dig Dis 2019; 37(6): 509–517. doi: 10.1159/000500717.
77. Bhanji RA, Montano-Loza AJ, Watt KD. Sarcopenia in cirrhosis: looking beyond the skeletal muscle loss to see the systemic disease. Hepatology 2019; 70(6): 2193–2203. doi: 10.1002/hep.30686.
78. Grønbæk H, Rodgaard-Hansen S, Aagaard NK et al. The soluble macrophage activation markers sCD163 and mannose receptor (sMR) predict mortality in patients with liver cirrhosis without or with acute-on-chronic liver failure (ACLF). J Hepatol 2016; 64(4): 813–822. doi: 10.1016/j.jhep.2015.11.021.
79. Tan EX, Wang MX, Pang J et al. Plasma exchange in patients with acute and acute-on-chronic liver failure: a systematic review. World J Gastroenterol 2020; 26(2): 219–245. doi: 10.3748/wjg.v26.i2.219.
80. Gustot T, Fernandez J, Szabo G et al. Sepsis in alcohol-related liver disease. J Hepatol 2017; 67(5): 1031–1050. doi: 10.1016/j.jhep.2017.06.013.
81. Tritto G, Bechlis Z, Stadlbauer V et al. Evidence of neutrophil functional defect despite inflammation in stable cirrhosis. J Hepatol 2011; 55(3): 574–581. doi: 10.1016/j.jhep.2010.11.034.
82. Xu R, Bao C, Huang H et al. Low expression of CXCR1/2 on neutrophils predicts poor survival in patients with hepatitis B virus-related acute-on-chronic liver failure. Sci Rep 2016; 6: 38714. doi: 10.1038/srep38714.
83. Artru F, Bou Saleh M, Maggiotto F et al. IL-33/ST2 pathway regulates neutrophil migration and predicts outcome in patients with severe alcoholic hepatitis. J Hepatol 2020; 72(6): 1052–1061. doi: 10.1016/j.jhep.2019.12.017.
84. Rolas L, Makhezer N, Hadjoudj S et al. Inhibition of mammalian target of rapamycin aggravate the respiratory burst defect of neutrophils from decompensated patients with cirrhosis. Hepatology 2013; 57(3): 1163–1171. doi: 10.1002/hep.26109.
85. Rolas L, Boussif A, Weiss E et al. NADPH oxidase depletion in neutrophils from patients with cirrhosis and restoration via toll-like receptor 7/8 activation. Gut 2018; 67(8): 1505–1516. doi: 10.1136/gutjnl-2016-313443.
86. Papayannopoulos V. Neutrophil extracellular traps in immunity and disease. Nat Rev Immunol 2018; 18(2): 134–147. doi: 10.1038/nri.2017.105.
87. Agraz-Cibrian JM, Segura-Ortega JE, Delgado-Rizo V et al. Alterations in neutrophil extracellular traps is associated with the degree of decompensation of liver cirrhosis. J Infect Dev Ctries 2016; 10(5): 512–517. doi: 10.3855/jidc.7165.
88. Bukong TN, Cho Y, Iracheta-Vellve A et al. Abnormal neutrophil traps and impaired efferocytosis contribute to liver injury and sepsis severity after binge alcohol use. J Hepatol 2018; 69(5): 1145–1154. doi: 10.1016/j.jhep.2018.07.005.
89. Rajkovic IA, Williams R. Abnormalities of neutrophil phagocytosis, intracellular killing and metabolic activity in alcoholic cirrhosis and hepatitis. Hepatology 1986; 6(2): 252–262. doi: 10.1002/hep.1840060217.
90. Gomez F, Ruiz P, Schreiber AD. Impaired function of macrophage Fc receptors and bacterial infection in alcoholic cirrhosis. N Engl J Med 1994; 331(17): 1122–1128. doi: 10.1056/NEJM199410273311704.
91. Korf H, du Plessis J, van Pelt J et al. Inhibition of glutamine synthetase in monocytes from patients with acute-on-chronic liver failure resuscitates their antibacterial and inflammatory capacity. Gut 2019; 68(10): 1872–1883. doi: 10.1136/gutjnl-2018-316888.
92. Brenig R, Pop OT, Triantafyllou E et al. Expression of AXL receptor tyrosine kinase relates to monocyte dysfunction and severity of cirrhosis. Life Sci Alliance 2020; 3(1): e201900465. doi: 10.26508/lsa.201900465.
93. Bernsmeier C, Pop OT, Singanayagam A et al. Patients with acute-on-chronic liver failure have increased numbers of regulatory immune cells expressing the receptor tyrosine kinase MERTK. Gastroenterology 2015; 148(3): 603–615. doi: 10.1053/j.gastro.2014.11.045.
94. Triantafyllou E, Woollard KJ, McPhail MJW et al. The role of monocytes and macrophages in acute and acute-on-chronic liver failure. Front Immunol 2018; 9: 2948. doi: 10.3389/fimmu.2018.02948.
95. Singanayagam A, Triantafyllou E. Macrophages in chronic liver failure: diversity, plasticity and therapeutic targeting. Front Immunol 2021; 12: 661182. doi: 10.3389/fimmu.2021.661182.
96. Riva A, Mehta G. Regulation of monocyte-macrophage responses in cirrhosis – role of innate immune programming and checkpoint receptors. Front Immunol 2019; 10: 167. doi: 10.3389/fimmu.2019.00167.
97. Lario M, Muňoz L, Ubeda M et al. Defective thymopoiesis and poor peripheral homeostatic replenishment of T-helper cells cause T-cell lymphopenia in cirrhosis. J Hepatol 2013; 59(4): 723–730. doi: 10.1016/j.jhep.2013.05.042.
98. Noor MT, Manoria P. Immune dysfunction in cirrhosis. J Clin Transl Hepatol 2017; 5(1): 50–58. doi: 10.14218/JCTH.2016.00056.
99. Riva A, Patel V, Kurioka A et al. Mucosa-associated invariant T cells link intestinal immunity with antibacterial immune defects in alcoholic liver disease. Gut 2018; 67(5): 918–930. doi: 10.1136/gutjnl-2017-314458.
100. De Biasi S, Gibellini L, Lo Tartaro D et al. Circulating mucosal-associated invariant T cells identify patients responding to anti-PD-1 therapy. Nat Commun 2021; 12(1): 1669. doi: 10.1038/s41467-021-21928-4.
101. Lebossé F, Gudd C, Tunc E et al. CD8+ T cells from patients with cirrhosis display a phenotype that may contribute to cirrhosis-associated immune dysfunction. EBioMedicine 2019; 49: 258–268. doi: 10.1016/j.ebiom.2019.10.011.
102. Peter J, Frey O, Stallmach A et al. Attenuated antigen-specific T cell responses in cirrhosis are accompanied by elevated serum interleukin-10 levels and down-regulation of HLA-DR on monocytes. BMC Gastroenterol 2013; 13: 37. doi: 10.1186/1471-230X-13-37.
103. Doi H, Iyer TK, Carpenter E et al. Dysfunctional B-cell activation in cirrhosis resulting from hepatitis C infection associated with disappearance of CD27-positive B-cell population. Hepatology 2012; 55(3): 709–719. doi: 10.1002/hep.24689.
104. Laso FJ, Madruga JI, Girón JA et al. Decreased natural killer cytotoxic activity in chronic alcoholism is associated with alcohol liver disease but not active ethanol consumption. Hepatology 1997; 25(5): 1096–1100. doi: 10.1002/hep.510250508.
105. Bernsmeier C, Triantafyllou E, Brenig R et al. CD14+ CD15- HLA-DR- myeloid-derived suppressor cells impair antimicrobial responses in patients with acute-on-chronic liver failure. Gut 2018; 67(6): 1155–1167. doi: 10.1136/gutjnl-2017-314 184.
106. Oberholzer A, Oberholzer C, Moldawer LL. Interleukin-10: a complex role in the pathogenesis of sepsis syndromes and its potential as an antiinflammatory drug. Crit Care Med 2002; 30(Suppl 1): S58–S63.
107. Peter J, Frey O, Stallmach A et al. Attenuated antigen-specific T cell responses in cirrhosis are accompanied by elevated serum interleukin-10 levels and down-regulation of HLA-DR on monocytes. BMC Gastroenterol 2013; 13: 37. doi: 10.1186/1471-230X-13-37.
108. Markwick LJ, Riva A, Ryan JM et al. Blockade of PD1 and TIM3 restores innate and adaptive immunity in patients with acute alcoholic hepatitis. Gastroenterology 2015; 148(3): 590–602.e10. doi: 10.1053/j.gastro.2014.11.041.
109. Rimola A, Soto R, Bory F et al. Reticuloendothelial system phagocytic activity in cirrhosis and its relation to bacterial infections and prognosis. Hepatology 1984; 4(1): 53–58. doi: 10.1002/hep.1840040109.
110. Sander LE, Sackett SD, Dierssen U et al. Hepatic acute-phase proteins control innate immune responses during infection by promoting myeloid-derived suppressor cell function. J Exp Med 2010; 207(7): 1453–1464. doi: 10.1084/jem.20091474.
111. Maini AA, Becares N, China L et al. Monocyte dysfunction in decompensated cirrhosis is mediated by the prostaglandin E2-EP4 pathway. JHEP Rep 2021; 3(6): 100332. doi: 10.1016/j.jhepr.2021.100332.
112. O‘Brien A, Fullerton J, Massey K et al. Immunosuppression in acutely decompensated cirrhosis is mediated by prostaglandin E2. Nat Med 2014; 20(5): 518–523. doi: 10.1038/nm.3516.
113. Zhou Q, Shi Y, Chen C et al. A narrative review of the roles of indoleamine 2,3-dioxygenase and tryptophan-2,3-dioxygenase in liver diseases. Ann Transl Med 2021; 9(2): 174. doi: 10.21037/atm-20-3594.
114. Van der Merwe S, Chokshi S, Bernsmeier C et al. The multifactorial mechanisms of bacterial infection in decompensated cirrhosis. J Hepatol 2021; 75(Suppl 1): S82–S100. doi: 10.1016/j.jhep.2020.11.029.
115. Villanueva C. Bacterial infections in patients with compensated cirrhosis and clinically significant portal hypertension: implications on the risk of developing decompensation and on survival. Hepatology 2019; 70: 36A–37A.
116. Fernández J, Acevedo J, Wiest R et al. Bacterial and fungal infections in acute-on-chronic liver failure: prevalence, characteristics and impact on prognosis. Gut 2018; 67(10): 1870–1880. doi: 10.1136/gutjnl-2017-314240.